









 はじめに 植物の形づくりの機構は特にシロイヌナズナやイネの突然変異体を手がかりとして明らかになってきましたが、これらの比較的単純な体制とゲノムを持つモデル植物を用いた研究だけでは全てを理解することができません。そのため、世界各地で、様々なモデル植物をもちいた研究が進展しています。 アサガオは花の色や模様だけでなく、形に関する変異体も豊富で、活性の高いトランスポゾン(動く遺伝子)があるというのも特徴の一つです。日本で大正~昭和初期にかけて行われた、たくさんの古典遺伝学的研究の成果も利用できます。
当研究室はこのアサガオの突然変異系統を1000系統以上保存している世界で唯一の機関で、文部科学省の主要な生物遺伝資源の整備事業、「ナショナルバイオリソースプロジェクト」の中核機関にも指定されています。そのため、モデル植物としての質を高めるための突然変異系統の収集・保存・解析、連鎖地図の作成等も行っています。 |
 ここに展示している植物は、どれも、れっきとしたアサガオです。形はアサガオとは似ても似つかなくても、遺伝子から見るとほんの数個の遺伝子が違っているだけで、普通のアサガオとほとんど差がありません。つまり、形のおかしくなったアサガオと正常なアサガオの遺伝子を比べて、違っている部分を調べる(クローニング)ことで植物の形づくりの仕組みを理解するために研究を行っています。また、これまで取られたアサガオの変異遺伝子のほとんどにトランスポゾンが挿入してたので、そのトランスポゾンの構造や転移機構の研究も行っています。
ここに展示している植物は、どれも、れっきとしたアサガオです。形はアサガオとは似ても似つかなくても、遺伝子から見るとほんの数個の遺伝子が違っているだけで、普通のアサガオとほとんど差がありません。つまり、形のおかしくなったアサガオと正常なアサガオの遺伝子を比べて、違っている部分を調べる(クローニング)ことで植物の形づくりの仕組みを理解するために研究を行っています。また、これまで取られたアサガオの変異遺伝子のほとんどにトランスポゾンが挿入してたので、そのトランスポゾンの構造や転移機構の研究も行っています。 今から1200年ほど前の奈良時代に薬草として日本に渡来したアサガオは江戸時代までは、原種の青花と、色変わりの白花のしかなかったのようです。江戸時代後期(文化文政期;1804~)に、たくさんの変わりもの(突然変異体)があらわれました。
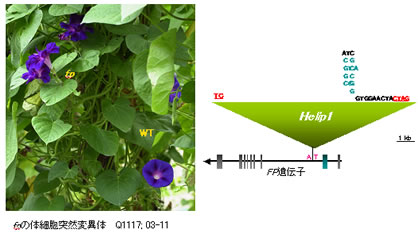
今から1200年ほど前の奈良時代に薬草として日本に渡来したアサガオは江戸時代までは、原種の青花と、色変わりの白花のしかなかったのようです。江戸時代後期(文化文政期;1804~)に、たくさんの変わりもの(突然変異体)があらわれました。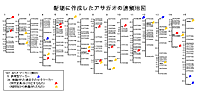


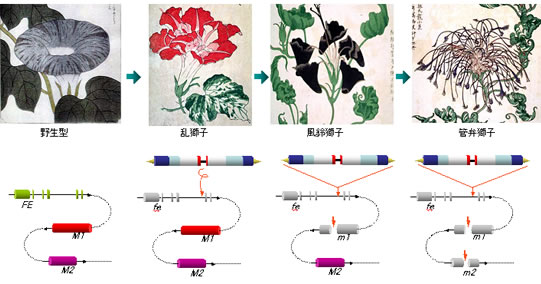
形態形成突然変異の研究(1):獅子(fe)遺伝子 
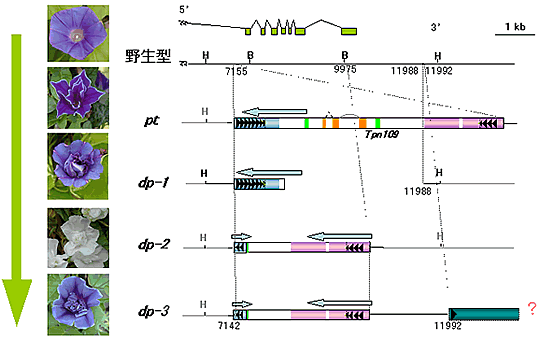
八重咲(pt)と牡丹(dp)変異は、花弁を増やし、花を豪華にするため、多くのアサガオ系統が持っている花のホメオティック変異です。種子の出来ない牡丹変異を最初に保存することがどうして可能だったのか、今までは謎に包まれていました。遺伝子解析によって、実は八重咲と牡丹変異は同じ遺伝子の突然変異で、まず種子のできる八重咲変異が起こって、それからトランスポゾンの転移によって、DP遺伝子のエクソンを欠失した、より表現型の強い牡丹変異が出てきたことがわかりました。 |
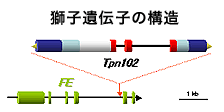 トランスポゾンの挿入を指標にして、獅子遺伝子のクローニングに成功しました。原因遺伝子はやはり背軸(裏)側の器官形成に関わるシロイヌナズナのKAN1と最も相同性の高い遺伝子でした。
トランスポゾンの挿入を指標にして、獅子遺伝子のクローニングに成功しました。原因遺伝子はやはり背軸(裏)側の器官形成に関わるシロイヌナズナのKAN1と最も相同性の高い遺伝子でした。Fatal error: Uncaught Error: Failed opening required 'footer.tmpl' (include_path='.:/usr/local/php/8.3/lib/php') in /home/opack/www/fair/a/a_6.html:127 Stack trace: #0 {main} thrown in /home/opack/www/fair/a/a_6.html on line 127